Journal club 2019
・Wever et al. A dual role for PP1 in shaping the Msn2-dependent transcriptional response to glucose starvation. EMBO J.(2005) 24,4115-4123
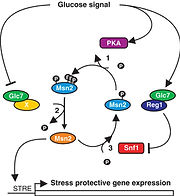
酵母では、グルコース枯渇によってストレス関連遺伝子の転写が迅速に誘導される。Msn2は、このストレス応答を調節する主な転写活性化因子であり、Msn2の核局在化とDNA結合はcAMP依存性プロテインキナーゼ(PKA)によって負に制御されている(Görner et al, 1998)。本研究において、急激なグルコース枯渇によるMsn2の活性化は、Msn2核移行シグナル(NLS)上の一過的な脱リン酸化と関係していることを発見した。このことから、プロテインホスファターゼ1(PP1)は、Msn2のNLSにおけるPKA依存性リン酸化のアンタゴニストであり、Msn2を標的とするグルコース枯渇シグナルのメディエーターの役割を持つことが見出された。また、タンパク質キナーゼSnf1はMsn2リン酸化部位の1つ(S582)を特異的に修飾し、Msn2を不活性化することから、Glc7(PP1の触媒サブユニット)の調節因子であるReg1に依存したPP1は、Snf1を抑制することにより、間接的にMsn2を活性化させることも明らかとなった。したがって、PP1のさまざまな活動は、急性グルコース枯渇時の転写段階でのストレス応答と適応の誘導に関与していると考えられる。 (紹介者 今井)
・Meitinger et al.(2014) A memory system of negative polarity cues prevents replicative aging. Cell. 2014 Nov 20;159(5):1056-1069. doi: 10.1016/j.cell.2014.10.014. Epub 2014 Nov 13.
Rho-GTPaseであるCdc42は細胞極性化のマスターレギュレーターとして真核生物で高く保存されており、酵母細胞はG1期に出芽する部位に極性を生じさせる。出芽した部位には痕跡としてCRMs(Cytokinesis ReMnants=bud scars)が母細胞上に残される。本研究では、著者らは酵母細胞がCRMs上に極性が再構築されないメカニズムを研究した。スカフォールドタンパク質のGps1は細胞分裂部位でCdc42依存的なCla4(出芽に関するキナーゼ)の活性化を阻害することが知られていた。Gps1はNap1,Nba1,Nis1と相互作用を示し細胞分裂部位に局在するが、Nba1とNis1はCRMsにも局在が見られた。Nba1はCdc42の阻害に関与しており、Rsr1によるCdc24の活性化を阻害している。また、膜貫通タンパク質のRax1-Rax2複合体がNba1とNis1をCRMsにアンカーし続けることによって、CRMsで再出芽することを抑制していた。また、CRMs上で出芽を繰り返すrga1Δnba1Δ株などではbud neckが細くなり、核分離の異常や細胞寿命の短縮化が引き起こされることも確認された。(紹介者 清水)
・Gomar-Alba et al.(2012) The Saccharomyces cerevisiae Hot1p regulated gene YHR087W (HGI1) has a role in translation upon high glucose concentration stress. BMC Mol Biol. 2012 Jun21;13;19.doi: 10.1186/1471-2199-13-19.
酵母は高濃度グルコース条件下で浸透圧ストレスを受けると、その応答としてHOG経路のHOG1のリン酸化によって開始される転写が制御される。この時、誘導される遺伝子の一つがYHR087W(HGI1)である。本研究では、YHR087Wの遺伝子の機能解析が行われた。解析の結果、YHR087Wは高濃度グルコースストレス下では、Sko1やHot1などの転写因子がHog1依存的にYHR087Wのプロモーター領域に結合することで転写が活性化されることが示唆された。また、YHR087Wpは翻訳開始因子との物理的な相互作用があることが分かり、YHR087W遺伝子の欠損と翻訳開始因子のelF4EであるCdc33の変異の組合せ(Δyhr087w cdc33ts)でハイグロマイシンに対する感受性の上昇が相乗的に見られ、更にYHR087W欠損は細胞の翻訳活性の低下を引き起こすことが明らかとなった。YHR087Wの欠損は遺伝子の翻訳効率に関わるポリソームとモノソームの比率に対しても影響し、著者らはこの影響が重要であると考えている。この論文では、YHR087Wの転写・翻訳の制御からその働きまで広く扱われており、これら結果をもとにより掘り下げた内容の研究が行われることが期待される。(紹介者 清水)
・Li et al.(2016) The mRNA cap-binding protein Cbc1 is required for high and timely expression of genes by promoting the accumulation of gene-specific activators at promoters. Biochim. Biophys.Acta,1859,405-419
著者らは、以前にcbc1Δ株では野生株と比べて浸透圧ストレス誘導性遺伝子の転写活性化が遅れることを報告している(Garre et al. Mol Biol Cell.2012)。本研究では、この現象が特定の遺伝子で起こるのか一般的に起こるのかを検証するとともに、浸透圧ストレス応答性遺伝子へのRNA polymeraseⅡのリクルートの遅れる原因についてHog1に依存する転写因子を用いて解析を行った。本研究結果から、転写活性化の遅れはほとんどの遺伝子で起こっており、Cbc1は浸透圧誘導性遺伝子の急速な転写活性化やHog1-Hot1複合体およびPICが効率的に浸透圧誘導性遺伝子のプロモーターに結合する上で必要であることを報告している。また、野生株とcbc1Δ株でTR(transcription rate)を比較した結果、、転写レベルが高い遺伝子については、発現レベルを高く維持するのにCbc1が必要であり、様々な環境ストレスに応答した遺伝子発現の調節にもCbc1が重要な役割を担う可能性があることも報告している。 (紹介者 今井)
・Gao et al.(2005)Cap-binding protein 1-mediated and eukaryotic translation initiation factor 4E-mediated pioneer rounds of translaion in yeast. Proc. Natl. Acad.Sci. USA.,102,4258-4263.
哺乳動物では、翻訳開始因子であるeIF4に結合したmRNAはNMDのターゲットにはならないが(Lejeune et al.(2002)EMBO J.,21,3536-3545)、酵母ではキャップ結合タンパク質であるCbc1pに結合したmRNAとeIF4Eに結合したmRNAのどちらもNMDのターゲットになることを報告している。著者らは図のようなS.cerevisiaeのmRNA翻訳モデルを提唱している。核内では、転写と並行してCbc2pとヘテロダイマーとして結合するCbc1pまたはeIF4Eが5'キャップに結合したmRNAが形成され、細胞質で両者とも翻訳のパイオニアラウンドを受ける。Cbc1pに結合したmRNAは、パイオニアラウンド中または後にeIF4Eに結合したmRNAに置換される。将来の研究は、Cbc1p結合mRNA翻訳の性質とNMDに対する重要性が明らかになると期待を寄せている。(紹介者 今井)
・Lien PTK et al.(2019) Pop2 phosphorylation at S39 contributes to the glucose repression of stress response genes, HSP12 and HSP26. PLoS ONE 14(4): e0215064.
出芽酵母のPop2タンパク質は遺伝子発現のレギュレーターとしての役割をもつCcr4-NOT複合体を構成するエキソヌクレアーゼで、転写後にmRNAのpoly A tail を分解し短くすることで遺伝子発現を調節する。これまでの研究で、Pop2のトレオニン97(T97)がリン酸化されることは知られていたが、そのリン酸化の生理的意義は不明である。本研究で、著者らはPop2がグルコース存在下でセリン39(S39)がリン酸化され、これがheat shock proteinであるHSP12やHSP26のmRNAレベルの制御に関与することを発見した。このリン酸化はHSP12やHSP26の発現を抑制するグルコースリプレッションとして機能していると考えられるが、転写段階に関与するのかmRNAの分解効率に関与するのかについては十分な検討が行われていない。Pop2のS39のリン酸化がどのようにHSP12とHSP26のmRNAレベルを制御するのか、その分子メカニズムの解明が期待される。(紹介者 清水)
・Luethy et al. (2019) Anesthetic pretreatment confers thermotolerance on Saccharomyces cerevisiae yeast. Biochem. Biophys. Res. Commun., doi: 10.1016/j.bbrc.著者らはAPC (Anesthetic preconditioning)と呼ばれる現象を解明するために、出芽酵母を用いて麻酔薬の一つであるTBE (Tribromoethanol)が熱耐性賦与に与える影響を解析した。APCとは麻酔薬(イソフルランやTBEなどのハロゲン化化合物)の投与によって虚血性障害から臓器が保護される現象のことで、詳しいメカニズムはわかっていない。今回の論文ではTBEで細胞を前処理する事により出芽酵母の熱耐性が上昇する事、Trp/Leuを多く含む培地や対数増殖期の細胞では熱耐性が上昇しにくいことなどを明らかにしている。APC研究に酵母をモデル生物として扱う有用性を示した論文であり、今後のAPCに関する研究の発展が期待される。(紹介者 吉田)
・Elena Garre et al.,Yeast mRNA cap-binding protein Cbc1/Sto1 is necessary for the rapid reprogramming of translation after hyperosmotic shock,Mol Biol Cell.2012 Jan 1;23(1):137-150.ハウスキーピング遺伝子をコードするmRNAは、ストレスを受けると翻訳が抑制され、P bodyやSG(stress granules)へと移動する。一方で、ストレス保護タンパク質は、細胞が生存できるようにストレス下でも翻訳が促進される。(Yamasaki and Anderson, Curr. Opin.Cell Biol., 2008; Spriggs et al. Mol. Cell, 2010)。その後、ストレスの適応した際にはP bodyやSGに保持されていたmRNAの一部が細胞質に戻り翻訳が再開される。このようなストレスに対する翻訳のリプログラミングが正しく行われない場合、癌や糖尿病などの病気につながると報告されている(Keene, Nat. Rev. Genet.,2007)。今日まで、酵母の浸透圧ストレス条件下における特定の遺伝子が翻訳されるメカニズムや関連因子は明らかになっていない。著者らは、浸透圧ストレスに応答する遺伝子の発現調節に関与する因子の一つとしてキャップ結合タンパク質であるCbc1を特定した。ポリソーム解析とGFPを用いたP body形成の顕微鏡観察から、Cbc1は浸透圧ストレス応答性のmRNAの翻訳に関与していることが明らかとなった。また、翻訳開始の段階でキャップ構造を認識する翻訳開始因子eIF4Eの温度感受性変異株cdc33tsを用いた実験では、浸透圧ストレス下でCbc1はeIF4Eの代わりの働きをすることが示唆された。これらの結果より、著者らは、浸透圧ストレス保護タンパク質をコードするmRNAがCbc1と核内で結合し、細胞質で翻訳されるモデルを提唱した。今後の課題としては、Cbc1を介した翻訳機構の全容解明と浸透圧以外のストレスでのCbc1の関与を検討していくことが挙げられる。(紹介者 今井)
・Kim et al.(2018) Synergy between the small intrinsically disordered protein Hsp12 and trehalose sustain viability after severe desiccation.eLife doi: 10.7554/eLife.38337.Anhydrobioteは著しい水の欠如に耐性を持つ微生物、植物、動物を指す。乾燥耐性の分子的メカニズムを理解することはストレスに関する生物学や乾燥耐性を持った作物を作成する方法に新たな知見を与えるかもしれない。著者らは、anhydrobioteである出芽酵母を用いて、トレハロースとhydrophilinであるHsp12が致死的な乾燥ストレス条件下でどのような働きを示すのか検証した。トレハロース合成酵素であるTps1とHsp12の二重欠損株は単独欠損株tps1Δやhsp12Δよりも乾燥ストレス感受性が顕著に上昇しており、この両者の因子に相乗効果が見られた。また、乾燥ストレス条件下のタンパク質の活性や凝集などのタンパク質恒常性についても検証した結果、hsp12Δとtps1Δに相乗効果が見られた。Hsp12は膜のリモデリング活性を持ち、リポソームをナノリポソームに分解する働きを持つ。また、線虫を用いて行われた実験でトレハロースは膜に介入することが以前に報告されている(Crowe et all., 1984,1987;Leslie et all., 1994)。これらの結果から、著者らはトレハロースの膜への介入によって乾燥誘導性の膜の損傷から保護し、損傷が生じれば、トレハロースはHsp12のリモデリング活性によって除去されるというモデルを提唱している。これらの因子が他の生物に乾燥耐性を与えるのか、研究が期待される。(紹介者 清水)
・Snober et al. (2009) Ssd1 Is Required for Thermotolerance and Hsp104-Mediated Protein Disaggregation in Saccharomyces cerevisiae. Mol. Cell. Biol., 29(1):187-200.酵母にとって50℃というのは致死的な熱ストレスだが、37℃という比較的弱い熱ストレスを経験することによって生育が可能になる。この適応現象は37℃の熱ストレス時にHSP(Heat Shock Protein)の発現量や、タンパク質を変性から保護する機能を持つトレハロースの合成量が増加する事などによって起こると考えられているが、詳しいメカニズムは明らかになっていない。今回著者らは熱ストレスへの適応に必要な遺伝子としてSSD1を新たに見出し、ストレス下での機能解析を行った結果、Ssd1はHsp104を介して変性タンパク質を処理していることが明らかになった。具体的には、37˚Cで30 min、その後46˚Cで30 min処理した際にSsd1がHsp104の6量体形成を促進していると考えられ、適応能力を欠くssd1Δ株ではHsp104が6量体を形成せずにSti1(Hsp70やHsp90といった分子シャペロンのATPase活性を調節することが知られている。)と相互作用している様子が観察された。また、ssd1Δ株ではHsp104が十分に発現しているにも関わらず、変性タンパク質の処理能力が低下している事をホタルルシフェラーゼ活性の回復実験で確認している。Ssd1は翻訳抑制機能を持つmRNA結合タンパク質でTOR経路やRAMネットワークなど様々な経路に関与していることが知られている。具体的な制御機構はまだわかっていないが、Ssd1によるHsp104の活性の制御がシビアな熱ストレスへの適応に必要であると考えられる。(紹介者 吉田)
・Jansen et al. (2009) Cbk1 regulation of the RNA-binding protein Ssd1 integrates cell fate with translational control. Curr. Biol., 19, 2114-2120.
RAMネットワークと呼ばれるシグナル伝達経路は酵母から後生動物まで高度に保存されており、酵母をモデルとしてよく研究されている。酵母では娘細胞の特異的遺伝子発現、細胞周期の調節、細胞分離、細胞壁の維持などさまざまな細胞プロセスをRAMネットワークで調節することが知られている。出芽酵母においてRAMネットワークの一部であるプロテインキナーゼCbk1は、通常、RNA結合タンパク質であり翻訳抑制因子でもあるSsd1をリン酸化することでSsd1を不活性化し、翻訳抑制を解除する。Ssd1は主にbud neckに局在しているので、Ssd1に結合しているSun4などの細胞壁の形態形成に関わるmRNAのpolarized localizationと翻訳を促す。対照的に、Cbk1のキナーゼ活性が阻害されると、Pボディとストレス顆粒にSsd1が局在化し、結合するmRNAの翻訳を抑制することが知られている。本研究では、高度に保存されている8つのSsd1リン酸化部位のすべてをアラニンまたはアスパラギン酸に置換した変異株(ssd1-8Aとssd1-8D)と野生株を比較したところ、Ssd1タンパク質に付加するリン酸基が減少し、特にssd1-8Aは致死性を示すことを見出している。また、Cbk1のキナーゼ活性が失われた細胞では、娘細胞の増殖が劇的に遅くなり、細胞壁溶解酵素に耐性を示す異常な細胞壁が形成されたことも併せて報告されている。Cbk1とSsd1の関係のような遺伝子発現の局所的な抑制は細胞壁を持たない真核生物でも見られる。例えば、後生動物では、プロテインキナーゼSrcとRNA結合タンパク質Zbp1の間でβ-アクチンの翻訳を調節している(Huttelmaier et al. Nature, 2005)。また、キイロショウジョウバエは、細胞の伸長と樹状突起の形態形成に不可欠なCbk1関連キナーゼをもっている(Cong et al., Development, 2001; Emoto et al., Cell, 2004; He et al., Mol. Biol. Cell, 2005)。著者らは、これら以外にも細胞プロセスに必要なシグナル伝達機構が新たに見つかるのではないかと期待を寄せている。 (紹介者 今井)
・Batool Ossareh-Nazari et al.(2014) Ubiquitylation by the Ltn1 E3 ligase protects 60S ribosomes from starvation-induced selective autophagy.J Cell Biol. 204(6): 909–917.
オートファジーの中で、リボソームサブユニットを選択的に分解する経路はリボファジーと呼ばれる。近年、酵母のリボファジーには脱ユビキチン化酵素が必要ということが報告された。この発見はE3リガーゼによるユビキチン化がリボファジーを阻害するのか、または脱ユビキチン化酵素によるユビキチンの分解がリボファジーの開始に必要なシグナルとなっているのか議論されていた。著者らは、Ltn1 E3 ligaseに着目してリボファジーにおけるリボソーム大サブユニットのユビキチン化の役割について検討した。窒素飢餓条件下でLtn1レベルはでは自己ユビキチン化し、分解されて減少することが観察された。一方、同条件下で脱ユビキチン化酵素のUbp3の減少は観察されなかったため、窒素飢餓条件下ではリボソーム大サブユニットの脱ユビキチン化が進むことでリボファジーが亢進すると考えられた。著者らは、非ストレス条件下ではリボソーム大サブユニットのLtn1によるユビキチン化によってリボファジーが阻害され、窒素飢餓条件下ではUbp3による脱ユビキチン化がリボファジー開始のシグナルとなるモデルを提唱している。(紹介者 清水)
・Sandra et al. (2014) Life-span extension by a metacaspase in the yeast Saccharomyces cerevisiae. Science, 344,1389-92.
PCD (programed cell death)の制御に必須のタンパク質として知られているカスパーゼは線虫から哺乳類までよく保存されたシステインプロテアーゼであり、単細胞生物である出芽酵母にはMCA1 (metacaspase1)というホモログ遺伝子が存在する。細胞を過酸化水素などで処理するとMca1依存的にPCDマーカーが現れることからMca1もPCDに関与していると考えられている。さらに、プロテオスタシスの異常によって形成されるタンパク質凝集体にMca1がリクルートされる事も確認されており、PCDだけでなくPQC (protein quality control)にもMca1が関与していると思われる。今回の実験で筆者らはHSP104とプロテアソームの存在下でMCA1の発現量を増加させると不良タンパク質及びその凝集体の蓄積が阻害され、さらに複製寿命が延びる事を新たに発見した。また、複製寿命の制御においてMca1のカスパーゼ活性とHsp40シャペロンであるYdj1の間に機能的な関連があることも明らかにしている。PQCとPCDに関与するMca1がどのような分子機構で複製寿命を制御しているのか、今後の研究が期待される。(紹介者 吉田)
・Mateljak et al. (2017) Evolved a-factor prepro-leaders for directed laccase evolution in Saccharomyces cerevisiae. Microb. Biotechnol. Nov;10(6):1830-1836.
真菌laccase(EC 1.10.3.2)は、フェノールや芳香族アミンなどの酸化を触媒し、同時に分子状酸素を水に還元する。laccaseは、有機物合成やバイオレメディエーションなど様々なバイオテクノロジー分野への応用が期待できるoxidaseのエリートである。異種タンパク質の発現や分泌には S. cerevisiae由来のα-ファクターのプレプロ配列 (a-factor-prepro leaders = α native )が古くから使用されている。α native は、19残基のアミノ酸からなるpre領域と3個のN結合型グリコシル化部位を有する64残基のアミノ酸のpro領域を含んでおり、通常、pre領域は、シグナルペプチダーゼの作用によって小胞体膜から除去され糖鎖修飾される。その後、α native 結合タンパク質はゴルジへと輸送され、さらにグリコシル化される。pro 領域はシャペロン様活性を示すと考えられており、Kex2、STE13プロテアーゼの作用を通してゴルジにおいてプロセシングされる。しかしながら、α native では異種宿主において真菌laccaseの発現や分泌が乏しいことから、本研究では、S. cerevisiaeにおける真菌laccaseの発現や分泌に対する、変異が導入されたα-ファクターのプレプロ配列(α PM1L, α PCL)の使用効果について検討した。まず S.cerevisiae由来の α native および α PM1L, α PCL と酸化還元電位の異なる4つのlaccase(PM1L, PcL, TspC30L, MtL)を融合した。それらのprepro-leader:laccase fusion(計12種)を形質転換した後、発現誘導物質としてガラクトースを用いた選択的発現培地中で増殖させ、その分泌および活性を評価した。その結果、α native よりも α PM1L, α PCLで、PcL、PM1LおよびTsp30Lの分泌が増強された。富栄養培地を含むフラスコ培養の場合、α native よりも α PM1L, α PCL で、結合したlaccaseの全ての分泌が増強された。α PM1L, α PCL には共通する変異箇所があり、それらは、pre領域の疎水性や、糖固定に対する親和性の変化に関与している可能性がある。また、α PM1L および α PCL にそれぞれ見いだされたpro領域における突然変異は、Ste3プロセシング部位を修飾し、最終成熟段階のゴルジ区画におけるKex2の性能に影響を及ぼすことが示唆された。これらの結果は、真菌laccaseの機能的発現のためにα PM1L, α PCL を使用することの重要性を示唆しており、それは酵母における異種タンパク質発現のためのユニバーサルシグナルペプチドを設計するための新たな道を開くと期待される。(紹介者 福田)
・Eisenman et al. (2012) Synthesis and assembly of fungal melanin. Microbiol. Biotechnol.,93,931-940. 自然界の至るところに見いだされるメラニンは、紫外線(UV)、酸化剤および電離放射線などの環境ストレスから細胞を保護する役割を持つ重要な色素である。多くの微生物がメラニンを生産するが、真菌のメラニンは、宿主の免疫応答に影響を及ぼし、病原性に寄与し得ることから、その構造や合成系の解明が精力的に研究されている。真菌のメラニン合成には2つの経路が存在する。多くの真菌は、1,8-ジヒドロキシナフタレンを介して内因性基質からメラニンを合成する(DHN経路)。この経路はポリケタイドシンターゼ(PKS)によって触媒される。一方、いくつかの真菌は、哺乳類のメラニン生合成に似た経路で、チロシンあるいはL-3,4-ジヒドロキシフェニルアラニン(L-DOPA)を初発の基質としてラッカーゼまたはチロシナーゼが関与するL-DOPA経路でメラニンを産生する。メラニンの化学構造は不明なものもあるが、一般的に粒状構造を有することが顕微鏡観察によって確認されている。真菌では、メラニン顆粒は細胞壁に局在し多糖類に架橋される可能性が高い。最近の研究は、真菌メラニンが哺乳類のメラノソームに似た内部小胞で合成され細胞壁に輸送されるケースもあることが報告されている。メラニン合成の場所を隔離することによって、有害な中間体から細胞内の様々な成分を保護していると考えられる。また、銅は、DHNおよびL-DOPA経路の両方で補酵素として重要である。メラニン合成経路や構造を解明することで、その放射線防護特性や種々の物質に結合する特性を生かした利用が広がることが期待される。(紹介者 福田)
・Christoph et al. (2019) Mitochondrial protein translocation-associated degradation. Nature, 569,679-683 ミトコンドリアタンパク質はTOM複合体 (translocase of the outer membrane complex) を介して細胞質側からミトコンドリアへと運び込まれる。この輸送機能に障害が発生するとプロテアソーム活性の増加 (UPRam : the unfolded protein response activated by the mistargeting of proteins) 、シャペロンの増加、ミトコンドリアの酸化的リン酸化抑制、細胞質タンパク質の生産量の調整 (mPOS : mitochondrial precursor over-accumulation stress) 、さらには外膜に蓄積した異常な前駆体たんぱく質のMsp1を介した除去の促進 (mitoCPR : mitochondrial compromised protein import response) といった様々なストレス応答が誘導されることが知られていた。しかし、非ストレス下においてTOM複合体の途中で詰まったタンパク質がどのような機構で処理されているのかは解明されていなかった。今回の実験で著者らは非ストレス下でTOM複合体に詰まった不良タンパク質がmitoTAD (mitochondrial protein translocation-associated degradation) と名付けられた経路で分解を受けることを新たに解明した。mitoTADでは、ERAD (endoplasmic reticulum-associated degradation) 関与因子であるUbx2がTOM複合体に詰まってしまった不良タンパク質を感知して、同じくERADに関与するCdc48複合体を誘導して、不良タンパク質を取り除いている。エネルギーの生産工場であるミトコンドリア機能の維持はアルコール発酵が可能な酵母といえども重要であり、その恒常的なメンテナンスを行う機構を特定したのは意義深いことである。また、ミトコンドリアのメンテンナンスはあらゆる真核生物において生存にかかわる重要な機構であると考えられるため、酵母に限らず様々な真核生物の研究を発展させうる重要な発見である。 (紹介者 吉田)
・Takahara and Maeda (2012) Transient sequestration of TORC1 into stress granules during heat stress. Mol. Cell, 47(2):242-252免疫抑制剤であるラパマイシンのターゲットとして知られているTORC1 (Target of rapamycin complex 1)はタンパク質合成や、リボソーム生合成、オートファジーなどに関与する重要なタンパク質複合体であり、様々なストレスによってその活性が抑制されることが知られていたが、詳細な分子機構は不明であった。本論文で著者らは、Pbp1の過剰発現がTORC1をSGs(ストレス顆粒)中に隔離する事でTORC1のシグナルを抑制する事を新たに明らかにした。さらに、このようなTORC1のSGsへの隔離がヒートショック下でも起こる事を見出し、加えてSGsのdisassemblyがヒートショックからのリカバリー時にTORC1の再活性化に直接関与している事を明らかにした。また、本来はTORC1によって活性化されるSch9を構成的に活性化する事でヒートショック下での変異の発生率が有意に上昇した事から、ヒートショック下でTORC1がSGsに隔離されて不活性化されることで細胞がDNA損傷から保護される可能性を述べている。このような調節機構は、ヒートショック下のみに限らず、その他の様々なストレス下でも機能していると考えられるため、これまで知られていなかったSGsがストレス応答へ関与するメカニズムがこの研究によって解明されていく事が期待される。(紹介者 吉田)
・Seda et al.(2015)TORC1 and TORC2 work together to regulate ribosomal protein S6 phosphorylation in Saccharomyces cerevisiae Mol. Biol. Cell, 27(2): 397–409. 真核生物のribosomal protein S6(Rps6)が栄養飢餓条件において脱リン酸化が引き起こされることは以前から知られているが、その機能は不明のままである。本論文では酵母におけるRps6のSer-232残基およびSer-233残基のリン酸化状態がTOR複合体1(TORC1)およびTORC2によって調節されていることを示している。TORC1はAGCファミリーキナーゼYpk3およびPP1ホスファターゼGlc7を介してSer-232残基およびSer-233残基のリン酸化を調節するのに対して、TORC2はYpk1/2を介してSer-232残基のみのリン酸化を調節することを明らかにした。また、 Rps6の非リン酸化変異体では生育が抑制され、タンパク質含有量の低下、40S リボソームサブユニットの合成阻害などの特徴が確認された。さらに、ポリソームプロファイリングとリボソームプロファイリングをおこなって、Rps6のリン酸化が翻訳に与える影響を検討したが、有意的な差は認められなかった。以上の結果から、著者らは酵母におけるRps6のリン酸化を調整するシグナル伝達カスケードを明らかにし、非リン酸化変異体における影響を示した。
・Ren et al. (2016) Identification of YPL014W (Cip1) as a novel negative regulator of cyclin-dependent kinase in Saccharomyces cerevisiae. Genes Cells,21(6):543-552. 出芽酵母ではCln1~3およびClb1~6が細胞周期に応じて発現量を変化させ、Cdk1(Cyclin dependent kinase 1)と複合体を形成して活性化する事によって細胞周期が進行する。この複合体はCKI( cyclin dependent kinase inhibitor)によって不活性化される事が知られており、出芽酵母のCKIとしてSic1やFar1などがすでに同定されている。著者らは今回の実験で新たにYPL014W(Cip1)がCKIとして機能する事を特定し、詳細な機能を調査した。論文中ではCip1の細胞周期に応じたリン酸化修飾、Cln2-Cdk1のリン酸化活性の阻害、過剰発現による細胞周期の停止、欠損時にG1/S移行が加速する事などについて述べている。最近になって機能が特定されたためCip1に関する論文はまだ少ないが、細胞周期において重要な機能を担っている可能性を秘めており、今後の研究が期待される。(紹介者 吉田)
・Guerra-Moreno et al. (2015) Proteomic Analysis Identifies Ribosome Reduction as an Effective Proteotoxic Stress Response. J. Biol. Chem., 290(50):29695-706. ストレス応答は潜在的に危険な脅威を識別し、それらを軽減する適応型細胞プログラムである。ミスフォールドタンパク質の蓄積は、複数のメカニズムを介して細胞に害を及ぼす可能性がある偏在的ストレスである。ヒ素などの三価メタロイドはタンパク質のミスフォールディングを引き起こす。ヒ素の毒性の分子的根拠は完全には解明されていないが、解糖系でのATP産生の阻害、ピルビン酸デヒドロゲナーゼの直接阻害、DNA損傷、酸化的損傷、キナーゼ阻害および分子シャペロンの阻害など様々な作用が報告されている。本研究では、ヒ素処理時の細胞応答を調べるために、TMT-MS分析によるプロテオミクス解析を行った。その結果、三価ヒ素が細胞内タンパク質を同化状態から異化状態へと広範な再編成をもたらすことを明らかにした(ヒ素が細胞内タンパク質の恒常性に関与)。具体的には、三価ヒ素で処理した細胞でタンパク質分解経路であるプロテアソームおよびオートファジーの構成因子 (Rpn4, Pre2, Rpn9, Cuz1, Atg8, Atg19, Atg33など) の発現量増加が確認された。一方、タンパク質合成を制御する経路の成分であるリボソーム自体は減少した。後者の観察は適応反応であり、ヒ素による有毒な作用ではないことは、ヒ素処理後のリボソーム成分欠損株(rpl20aΔ,rpl39Δなど)の生存率の向上によって示された。以上の結果から、三価ヒ素の主な細胞毒性が、新たに翻訳されたタンパク質のミスフォールドに由来し、翻訳活性の抑制が迅速で効果的かつ可逆的なProteotoxic Stress Responseであることを示した。(紹介者 福田)
・Koch et al.,(2012) Yar1 protects the ribosomal protein Rps3 from aggregation. J. Biol. Chem., 287(26): 21806-15. 酵母では毎分2000個のリボソームが合成されなければならない。そのため、リボソームタンパク質の迅速な産生やそれらの核への効率的な輸送、リボソームサブユニットへの正しい組み込みなどは最適な増殖速度を維持する為の必要条件である。40Sのリボソームサブユニットを構成しているタンパク質Rps3は正の電荷を帯びているため、核内に輸送されるまでに凝集を引き起こしやすいと考えられている。本論文ではアンキリンリピートタンパク質Yar1がRps3と直接相互作用し、Rps3の凝集を防ぎ、40Sのプレリボソームサブユニットが組み立てられるメカニズムを報告している。 yar1∆欠損株とrps3∆欠損株は野生型と比較すると生育の遅れを示した。欠損株では20SのrRNAが最も多く含まれていることから、20SrRNAから18SrRNAのプロセシングが阻害されることが原因であると考えられた。また、Yar1がin vivo、in vitroの両方においてRps3の凝集を抑制したことから、Yar1がRps3の可溶性を維持し、効率的なリボソームサブユニットの組み立てに関わっていると示唆された。これらのデータから著者らは、Yar1がRps3の特異的なシャペロンであると結論付けている。
・Kroschwald et al,(2018) Different Material States of Pub1 Condensates Define Distinct Modes of Stress Adaptation and Recovery. Cell Rep., 23(11);3327-3339
グルコースの枯渇やヒートショック(46˚C)などのシビアなストレスを受けると、酵母細胞ではSG(Stress Granule)が形成される。SGとはRNA及びRNA結合タンパク質(Pub1など)からなる顆粒で、生体膜を用いずに液-液相分離によって細胞質から分離されている。液-液相分離とは水溶性の物質どうしが水と油のように分離する現象のことである。今回の実験で使用されているPub1はグルコース枯渇またはヒートショックにより凝集体を形成する性質を持つpoly(A)+ RNA結合タンパク質であり、筆者らはPub1が形成する凝集体がストレスに応じて性質を変えることを新たに発見した。グルコース枯渇に伴って生じるpHの低下ではゲル状の凝集体を形成し、ヒートショック時には強固な凝集体を形成する。この反応がin vitroでも発生していることから、Pub1がストレスに応じて凝集体の性質を変化させることでストレスの種類を感知するバイオセンサーとして機能している可能性が示唆された。この結果は他のSG構成タンパク質もストレスに応じて性質を変化させる可能性を示しており、今後のSGに関する研究のさらなる発展が期待できる。(紹介者 吉田)
・Dhingra et al. (2018) Replication protein A (RPA) SUMOylation positively influences the DNA damage checkpoint response in yeast. J Biol Chem. in press
DNA損傷応答(DDR:DNA Damage Response)を誘導する経路のうち最も研究されているのはDDC(DNA Damage Checkpoint)である。DDCはDNAの損傷を認識して1本鎖になったDNAがRPA(Replication Protein A : Rfa1-3で構成される3量体)と結合する事で開始され、Mec1とその補因子を損傷部位に誘導する。こうして活性化されたMec1によりRad53がSgs1などを介して活性化される事で反応が進行し、細胞周期の停止及びDNAの損傷修復が誘導される。さらに、DDIS(DNA Damage Induced SUMOylation)もRPAがssDNAと結合する事によって誘導され、RPAがSUMO(Small Ubiquitin like Modifier)によって修飾される事で進行する事が知られている。DDCとDDISは並行して起こると考えられており、クロストークに関する研究はあまりされていなかった。今回の研究で筆者らは、SUMO化したRfa1がSgs1と相互作用する事でDDC経路の進行に関与する事を新たに発見した。他にも一部のエピトープタグをRfa1へ導入する事で細胞のDNA損傷誘導薬剤への感受性が高まるためSUMO化Rfa1の検出には適さない事を確認しており、実験系の最適化にも貢献している。今後の両経路のクロストークに関する研究のさらなる発展が期待される。(紹介者 吉田)